主な違い:フォトシステムIIは、フォトシステムIIより前に発見されたため、「I」と名付けられました。 しかしながら、光合成の過程において、光化学系IIは光化学系Iよりも先に作用する。両者の主な違いはそれらが反応する光の波長である。 光化学系Iは700nmより短い波長の光を吸収するが、光化学系IIは680nmより短い波長の光を吸収する。 しかしながら、それらは両方とも酸素光合成の過程において同様に重要である。

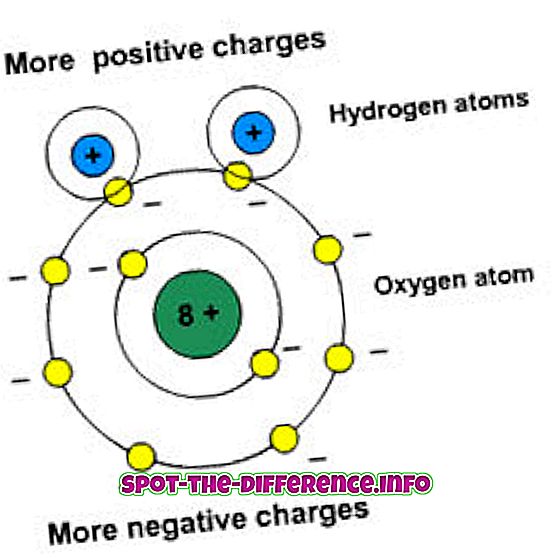
植物、藻類、そして多くの種類のバクテリアが光合成の過程で参加します。 それは植物や他のほとんどの種類のバクテリアの主要なエネルギー源の一つです。 植物とシアノバクテリアが酸素光合成を行うためには、それらは光化学系IとIIの両方を必要とする。 酸素性光合成は、二酸化炭素と水を使って酸素とエネルギーを作り出します。
光化学系は、光合成に関与するタンパク質複合体の構造単位です。 それらは、光合成の主要な光化学、すなわち光の吸収ならびにエネルギーおよび電子の伝達を実行する。 植物や藻類では、光化学系は葉緑体にありますが、光合成細菌では細胞質膜にあります。
それが光化学系IIの前に発見されたので、光化学系私は「私」と名付けられました。 しかしながら、光合成の過程において、光化学系IIは光化学系Iよりも先に作用する。両者の主な違いはそれらが反応する光の波長である。 光化学系Iは700nmより短い波長の光を吸収するが、光化学系IIは680nmより短い波長の光を吸収する。 しかしながら、それらは両方とも酸素光合成の過程において同様に重要である。
光化学系IはクロロフィルA分子P700を含み、これは700nmより短い波長を吸収する。 それは、そのアンテナシステム内の付随する補助顔料に加えて、そして光システムIIからの電子輸送チェーンから、光子からエネルギーを受け取る。 それは、NADP +(ニコチンアミドアデニンジヌクレオチドホスフェート)をNADPH + H +に還元するために、または単にプロトンポンプ(プラストキノン、またはPQ)に動力を与えるために光からのエネルギーを使用する。
光依存性の光合成における最初のタンパク質複合体である光化学系IIは、680 nmより短い波長の光を吸収するクロロフィルA分子P 680を含む。 それは、そのアンテナシステム内の光子および関連する補助顔料からエネルギーを受け取り、それを使用して水分子を酸化し、プロトン(H +)およびO 2を生成し、そして電子を電子輸送鎖に通過させる。

光合成の過程で、光化学系IIは光を吸収し、それを使用して反応中心クロロフィル中の電子がより高いエネルギーレベルに励起され、一次電子受容体によって捕捉される。 光化学系IIでは、4つのマンガンイオンのクラスターが水から電子を抽出し、それは次に酸化還元活性チロシンを介してクロロフィルに供給されます。
電子は次に光励起され、それはチラコイド膜中に設定された電子輸送鎖を通してチトクロームb 6 f錯体を通って光化学系Iに移動する。 電子のエネルギーは化学浸透と呼ばれるプロセスを通して利用されます。 このエネルギーは、ATPを生成するためのプロトン推進力を提供するために、膜を通って内腔へ水素(H +)を輸送するために使用される。 ATPシンターゼが管腔内に存在するプロトンを膜を介して支質に輸送すると、ATPが生成される。 プロトンはプラストキノンによって輸送されます。 電子が一度だけ通過する場合、その過程は非環式光リン酸化と呼ばれる。
電子が光化学系Iに到達した後、それは光化学系Iの反応中心クロロフィルを満たす。次いで電子は光励起され、光化学系Iの電子受容体分子に捕捉される。電子はPS Iの周りの循環電子輸送を通過し続ける。あるいはフェレドキシンを通過して酵素NADP +レダクターゼに達する。 電子および水素イオンはNADP +に付加されてNADPHを形成し、次いでそれはカルビンサイクルに輸送されてATPと共にグリセリン酸3−リン酸と反応してグリセルアルデヒド3−リン酸を形成する。 グリセルアルデヒド3-リン酸は、植物がさまざまな物質を作るために使用できる基本的な構成要素です。